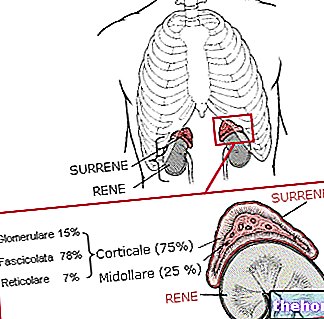
ASOCIĀCIJA (VAI "KONCENTRĀCIJA" VAI "SAITE")
Līdz šim mēs esam apstrādājuši Mendeļa dihibrīda (vai polihibrīda) krustojumu, pieņemot, ka dažādi alēliskie pāri faktiski ir atrodami dažādos homologos hromosomu pāros. Bet hromosomu pāru skaits, lai arī dažādās sugās ir atšķirīgs, atšķiras šaurās robežās (dažas sugas) sasniedz aptuveni simts hromosomu), savukārt gēnu skaitu var saskaitīt desmitos tūkstošu.
Laime, ka Mendela eksperimentiem izvēlētās rakstzīmes tika nošķirtas neatkarīgi (līdz ar to nesajaucot aprēķinus par fenotipu sadalījumu daudzhibrīda krusta F2). Ja blakus lokos tika atrasti divi alēļu pāri, likums tas tiktu saukts par asociācijas likumu.
Zinot, ka ļoti daudzām rakstzīmēm ir viena lokalizācija vienā hromosomā un ka tie ir hromosomu pāri, kas meiozē neatkarīgi nošķiras, var labi saprast, cik bieži gadās, ka divi rakstzīmju pāri, ja tie ir saistīti ar vecāku hromosomu vienādi arī gametā un līdz ar to arī ķermenī, pie kura tas nesīs savu ģenētisko materiālu.
Tādējādi mēs redzam, ka "asociācija ir" izņēmums, kas nebūt nav reti sastopams Mendela trešā likuma noteiktajā neatkarībā.
APMAIŅA VAI "PĀRTRAUKŠANA" UN REKOMBINĀCIJA
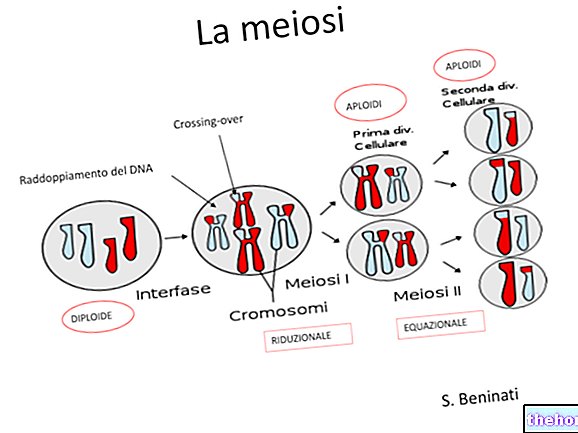
Runājot par mejozi, mēs esam norādījuši, ka pastāv divi dažādi ģenētiskā materiāla sajaukšanās momenti: viens ir hromosomu segregācija gametās, un to novēroja Mendela.
Otrs brīdis, kas faktiski ir pirms tam, ir tas, kurā katra homologo hromosomu pāra četras hromatīdas savstarpēji apmainās ar identiskām iezīmēm. Pēc šīs apmaiņas divi faktori, kas bija saistīti vienā hromosomā, tā vietā gametās būs neatkarīgi. tas, ka notiek apmaiņa, ir proporcionāls pirmajam tuvinājumam hromosomas garumam, un garākās hromosomās var būt pat vairāk nekā apmaiņa.
Šo parādību var noteikt citoloģiski, mikroskopā novērojot pietiekamu skaitu meiozes.
Atkārtotās atbilstības rādītājs ir ātrums, kādā jebkuras divas rakstzīmes, kas bija saistītas vecāku paaudzē, rekombinējas atšķirīgi F2.
Ja abi loki ir absolūti blakus, varbūtība, ka chiasms tos atdalīs, būs praktiski nulle. Rekombinācijas ātrums būs: n ° rekombinanti. Ja divi lokusi atrodas divās dažādās hromosomās, rekombinācijas ātrums būs 0,5 (vienāda varbūtība divām rakstzīmēm, kas pievienotas P paaudzei, nejauši atrasties kopā F2). Tāpēc rekombinācijas ātrums var svārstīties no 0,0 līdz 0,5. Nelieliem attālumiem hromosomā attālums un rekombinācijas ātrums ir tieši proporcionāli. Lielākiem attālumiem pastāv iespēja, ka starp diviem lokiem notiek divas apmaiņas. Tagad būs skaidrs, ka divi faktori, ko atdala divas apmaiņas, atkal ir saistīti. šajā brīdī ir skaidrs, ka tiek zaudēta proporcionalitāte starp lokusu attālumu un rekombinācijas varbūtību.
Vienā hromosomā saistītie loki veido “asociācijas grupas”. Ļoti attāliem lokiem var būt tāda varbūtība, ka tie tiks atdalīti apmaiņā, ka tie uzvedas kā neatkarīgi, taču katrs no tiem ar zemāku rekombinācijas ātrumu būs saistīts ar starpposma lokusiem.
Kad ir zināmi rekombinācijas ātrumi starp daudziem gēnu pāriem asociācijas grupā, var sākt veidot “ģenētiskās kartes”. Paturot prātā, ka attālumu starp diviem gēniem (a un b) izsaka rekombinācijas ātrums un ka a attālums no trešā gēna c var būt summa vai atšķirība attiecībā pret tā attālumu no b, ir iespējams rekonstruēt savstarpējo attālumu karti, kas būs šīs asociācijas grupas, tas ir, šīs hromosomas, ģenētiskā karte.
Tagad mums kopumā jāapsver daži jēdzieni, kas ierobežo genotipisko rakstzīmju fenotipisko izpausmi.
Vispirms mēs runāsim par iespiešanās un izteiksmīguma jēdzieniem, un pēc tam īpašu uzmanību veltīsim gēnu darbības regulēšanas parādībām.
PENETRANCE
Gēna iespiešanās atspoguļo tā spēju izpausties fenotipā. Caurlaidību mēra statistiski, saskaitot fenotipu biežumu, kas parāda šo raksturu no 100 genotipiem, kas to satur. Iezīme ar 0,7 penetranci ir iezīme, kas fenotipiski rodas 70% no tās genotipiskās biežuma.
EKSPRESIVITĀTE
Ekspresivitāte ir fenotipiskās izpausmes pakāpes kvantitatīvs novērtējums.
GĒNA DARBĪBAS REGULĒŠANA
Šūnas ražo visus fermentus un olbaltumvielas vienādā ātrumā un vienlaicīgi. Piemēram, Escherichia coli šūnas var apgādāt ar laktozes disaharīda enerģijas un oglekļa atomiem, jo tās spēj tās sadalīt glikozē un galaktozē, pateicoties beta-galaktozidāzes fermentam. Normālā E. coli, kurā var būt laktoze, ir aptuveni 3000 beta-galaktozidāzes molekulu, kas atbilst 3% šīs šūnas olbaltumvielu; ja nav laktozes, uz baktēriju šūnu būs tikai viena beta-galaktozidāzes molekula. Galaktosidāze tiks sintezēta no jaunām mRNS molekulām, kad to varēs izmantot. E. coli mutantu celmi, kas bagāti ar fermentu, ir zināmi pat tad, ja laktozes nav: šie mutanti ir nelabvēlīgā situācijā salīdzinājumā ar normālām šūnām, jo ir spiesti nevajadzīgi patērēt enerģiju un materiālus, lai ražotu fermentu, kas paliks bez substrāta. Vielas, kas izraisa fermentu daudzuma palielināšanos, kā tas ir laktozes gadījumā, tiks sauktas par induktoriem, bet fermenti - par inducējamiem. Citas vielas, arī tās īpašā veidā, izraisa noteiktu enzīmu ražošanu. Piemēram, arī E. coli, kas spēj veidot visas aminoskābes, kurās ir ogleklis un amonijs (NH3), konkrētas aminoskābes (piemēram, histidīna) klātbūtne barotnē bloķē visu enzīmu ražošanu pašas aminoskābes biosintēze: par šiem fermentiem tiks teikts, ka tie ir represējami. Baktēriju šūnās mRNS molekulas tiek nojauktas neilgi pēc to veidošanās, un tāpēc mRNS ražošanas kontrole nozīmē vienlaikus kontrolēt fermentatīvo sintēzi laiks.
OPERONS
Lai izskaidrotu, kā baktēriju šūna spēj kontrolēt savu fermentu ražošanu, Džeikobs un Monods formulēja operona hipotēzi; operonu veido vairāki gēni, kas ir funkcionāli saistīti un izkārtoti bez pārtraukuma gar DNS posmu.Operonu veido trīs dažādi gēnu veidi: promotors, kur sākas mRNS veidošanās; operators, kurā tiek veikta kontrole; viens vai vairāki strukturālie gēni, kas kodē fermentus vai citus proteīnus. Beta-galaktozidāzes sistēmā operons papildus beta-galaktozidāzes sistēmai ietver arī divus citus gēnus, kas strukturāli kodē citi fermenti, kas iesaistīti laktozes metabolismā.Šie gēni atrodas blakus viens otram un tiek pārrakstīti viens pēc otra pa to pašu DNS spirāli vienā mRNS molekulā.Šādi ražotās mRNS molekulas ir aktīvas ļoti īsu laiku, pēc tam tās iznīcina specifiski fermenti.
Savukārt operona darbību kontrolē cits gēns - regulators, kas var būt arī tālu no operona: šis regulators kodē proteīnu, ko sauc par represoru, kurš, šķiet, saistās ar DNS pie operatora gēna. starp promotoru un strukturālajiem gēniem faktiski bloķē mRNS ražošanu.
Represors savukārt tiek kontrolēts, un kontrole tiek veikta, izmantojot "signālvielu". Inducējamu enzīmu gadījumā šī viela ir "induktors". Induktors "saistās ar represoru molekulu, mainot tā formu tā, ka tā vairs nevar pielāgoties DNS: šajā gadījumā, tā kā starp promotoru un strukturālajiem gēniem nav represora. , represors var veidot mRNS molekulas un no tām olbaltumvielu molekulas. Līdz ar induktora piegādes izsīkumu regulators atkal atgūs kontroli, kas pārtrauks jaunu mRNS, tātad jaunu proteīnu, ražošanu. Beta-galaktozidāzes sistēmā induktors ir laktoze vai šai vielai ļoti līdzīga viela. pievienosies represoram, kas to deaktivizēs, lai nodrošinātu fermentu biosintēzi. Represējamu enzīmu gadījumā viela, kas darbojas kā "signāls", darbojas kā kodolpresors: represors ir aktīvs tikai tad, ja to apvieno ar kodolpresoru. Histidīna sistēmā, kurā ir iesaistīti duci dažādu enzīmu, šī ir aminoskābe kopā ar tās tRNS - kodorepresoru - histidīnu.
ALLOSTERISKĀ Mijiedarbība
Allosteriskā mijiedarbība, kas ietver fermenta inaktivāciju, mainot tā formu, nodrošina atšķirīgu veidu, kā regulēt šūnas metabolisko aktivitāti. Allosteriskā mijiedarbība ļauj precīzāk kontrolēt nekā operona induktora -represora sistēma, bet nesasniedz noderīgu rezultātu, ja tiek izslēgta konkrētas vielas biosintēze no pirmā posma - mRNS ražošanas.
KONTROLES SISTĒMAS EUCARIUS
Ir daži fakti, kas liek domāt, ka augiem un dzīvniekiem darbojas un dominē operonam līdzīga regulēšanas sistēma. Šo organismu hromosomas būtiski atšķiras no E. coli un citu prokariotu hromosomām. šūnas ir ļoti dažādas.Mitozes mehānisms ir tāds, ka katrai konkrētā auga vai dzīvnieka šūnai ir visa informācija
ģenētika, kas atrodas apaugļotā olšūnā. Tāpēc lielākā daļa gēnu jebkurā specializētā šūnā paliks neefektīvi visā šūnas dzīves laikā. Šo šūnu DNS vienmēr ir saistīta ar olbaltumvielām. Tāpēc ir iespējams, ka gēnu apspiešanai eikariotos ir nepieciešama tieši šī saistība starp DNS un olbaltumvielām.