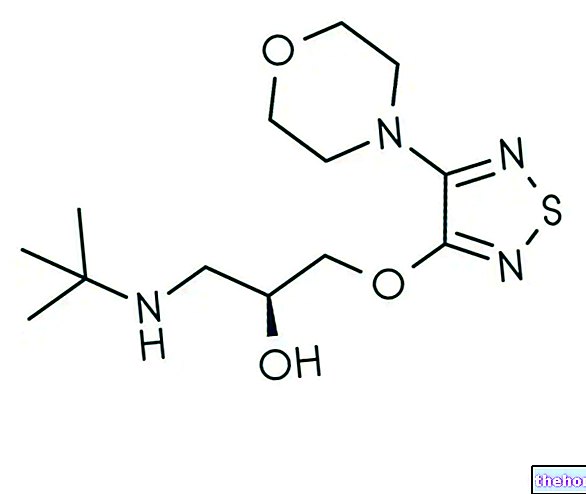
Olbaltumvielu elementārā analīze dod šādas vidējās vērtības: 55% oglekļa, 7% ūdeņraža un 16% slāpekļa; ir skaidrs, ka olbaltumvielas atšķiras viena no otras, bet to vidējais elementu sastāvs maz atšķiras no iepriekš norādītajām vērtībām .
Konstitucionāli proteīni ir makromolekulas, kas veidojas no dabīgām α-aminoskābēm; aminoskābes savienojas caur amīda saiti, kas tiek izveidota, reaģējot starp a-aminoskābes aminogrupu un citas a-aminoskābes karboksilgrupu.
Šo saiti (-CO-NH-) sauc arī par peptīdu saiti, jo tā saista peptīdus (aminoskābes kombinācijā):
iegūtais ir dipeptīds, jo tas sastāv no divām aminoskābēm. Tā kā dipeptīds vienā galā satur brīvu aminogrupu (NH2) un karboksilu otrā (COOH), tas var reaģēt ar vienu vai vairākām aminoskābēm un pagarināt ķēdi gan no labās, gan no kreisās puses ar tādu pašu reakciju redzams iepriekš.
Reakciju secība (kas, starp citu, nemaz nav tik vienkārša) var turpināties bezgalīgi: kamēr nav polimēra, ko sauc polipeptīds vai proteīns. Atšķirība starp peptīdiem un olbaltumvielām ir saistīta ar molekulmasu: parasti, ja molekulmasa ir lielāka par 10 000, mēs runājam par olbaltumvielām.
Aminoskābju saistīšana, lai iegūtu pat mazus proteīnus, ir grūts uzdevums, lai gan nesen tika izstrādāta automātiska olbaltumvielu ražošanas metode no aminoskābēm, kas dod izcilus rezultātus.
Tāpēc vienkāršākais proteīns sastāv no 2 aminoskābēm: saskaņā ar starptautisku vienošanos, pasūtītā aminoskābju numerācija olbaltumvielu struktūrā sākas no aminoskābes ar brīvo a-aminogrupu.
kodē šo proteīnu), kas saskaras ar nenozīmīgām ķīmiskām grūtībām.
Bija iespējams noteikt pasūtīto aminoskābju secību, sadaloties Edmanam: olbaltumviela reaģē ar fenilizotiocianātu (FITC); sākotnēji α-aminoskābju dubults uzbrūk fenilizotiocianātam, veidojot tiokarbamila atvasinājumu; pēc tam iegūtais produkts ciklizējas, iegūstot feniltio -hidantoīna atvasinājumu, kas ir fluorescējošs.
Edmans ir izstrādājis mašīnu, ko sauc par sekvenceri, kas automātiski pielāgo parametrus (laiku, reaģentus, pH utt.) Noārdīšanai un nodrošina olbaltumvielu primāro struktūru (par to viņš saņēma Nobela prēmiju).
Primārā struktūra nav pietiekama, lai pilnībā interpretētu olbaltumvielu molekulu īpašības; tiek uzskatīts, ka šīs īpašības būtībā ir atkarīgas no telpiskās konfigurācijas, ko olbaltumvielu molekulas mēdz pieņemt, saliekot dažādos veidos: tas ir, pieņemot to, kas ir definēts kā olbaltumvielu sekundārā struktūra.
Olbaltumvielu sekundārā struktūra mirgo, tas ir, tai ir tendence sadalīties karsējot; tad proteīni paši denaturējas, zaudējot daudzas raksturīgās īpašības. Papildus sildīšanai virs 70 ° C denaturāciju var izraisīt arī apstarošana vai reaģentu iedarbība (piemēram, no stiprām skābēm).
Olbaltumvielu denaturācija termiskā efekta dēļ tiek novērota, piemēram, karsējot olu baltumu: redzams, ka tas zaudē želatīna izskatu un pārvēršas par nešķīstošu baltu vielu. Tomēr proteīnu denaturācija noved pie to sekundārās struktūras iznīcināšanas, bet atstāj to primāro struktūru nemainīgu (dažādu aminoskābju savienošanu).
Olbaltumvielas iegūst terciāro struktūru, kad to ķēde, kaut arī joprojām ir elastīga, neraugoties uz sekundārās struktūras saliekšanos, salocās tā, lai radītu izliektu trīsdimensiju izkārtojumu cieta ķermeņa formā. Par terciāro struktūru ir atbildīgas galvenokārt disulfīda saites, kuras var izveidot starp cisteīnu -SH, kas izkliedēts pa molekulu.
No otras puses, ceturtā struktūra pieder tikai olbaltumvielām, ko veido divas vai vairākas apakšvienības. Piemēram, hemoglobīns sastāv no diviem olbaltumvielu pāriem (tas ir, visās četrās olbaltumvielu ķēdēs), kas atrodas tetraedra virsotnēs, lai radītu sfērisku struktūru; četras olbaltumvielu ķēdes satur kopā jonu un ne-kovalentās obligācijas.
Vēl viens kvartāra struktūras piemērs ir insulīna struktūra, kas, šķiet, sastāv no sešām olbaltumvielu apakšvienībām, kas sakārtotas pa pāriem trīsstūra virsotnēs, kuru centrā ir divi cinka atomi.
Šķiedru proteīni
Tie ir proteīni ar noteiktu stingrību un tiem ir daudz garāka asis nekā otram; šķiedrainais proteīns, kas dabā atrodas lielākā daudzumā, ir kolagēns (vai kolagēns).
Šķiedrains proteīns var uzņemties dažādas sekundārās struktūras: α-spirāli, β-loksni un, kolagēna gadījumā, trīskāršu spirāli; α-spirāle ir visstabilākā struktūra, kam seko β-lapa, bet vismazāk stabila no trim ir trīskāršā spirāle.
taisnība ja pēc galvenā skeleta (orientēts no apakšas uz augšu) tiek veikta kustība, kas līdzīga labās skrūves skrūvēšanai; kamēr spirāle ir no kreisā roka ja kustība ir līdzīga kreisās skrūves skrūvēšanai. Labās puses α-spirālēs aminoskābju -R aizvietotāji ir perpendikulāri proteīna galvenajai asij un ir vērsti uz āru, savukārt kreisajā pusē roku a -spirāles -R aizvietotāji ir vērsti uz iekšu. Labās puses spirāles ir stabilākas nekā kreisās, jo starp vati -R c "ir mazāk mijiedarbības un mazāk sterisku šķēršļu. Visas olbaltumvielās atrastās a-spirāles ir dekstrorotālas.
Α-spirāles struktūru stabilizē ūdeņraža saites (ūdeņraža tilti), kas veidojas starp katras aminoskābes karboksilgrupu (-C = O) un aminogrupu (-NH), kas vēlāk tika atrasta lineāra secība.
Piemērs proteīnam ar α-spirāles struktūru ir matu keratīns.
Pagarinot α-spirāles struktūru, tiek veikta pāreja no α-spirāles uz β-loksni; arī siltums vai mehāniskais spriegums ļauj pāriet no α-spirāles uz β-loksnes struktūru.
Parasti proteīnā β-lokšņu struktūras ir tuvu viena otrai, jo starp paša proteīna daļām var izveidot starpķēdes ūdeņraža saites.
Šķiedru proteīnos lielākā daļa olbaltumvielu struktūras ir sakārtota α-spirālē vai β-loksnē.
Globulāri proteīni
Tiem ir gandrīz sfēriska telpiskā struktūra (daudzo polipeptīdu ķēdes virziena izmaiņu dēļ); dažas būtnes daļas var izsekot līdz α-spirāles vai β-loksnes struktūrai, un citas daļas tā vietā nav attiecināmas uz šīm formām: izkārtojums nav nejaušs, bet organizēts un atkārtojas.
Līdz šim minētās olbaltumvielas ir pilnīgi viendabīgas struktūras vielas: tas ir, tīras kombinēto aminoskābju secības; šādi proteīni ir teikts vienkāršs; ir proteīni, kas sastāv no olbaltumvielu daļas un bezproteīna daļas (prostatas grupas), ko sauc par proteīniem konjugēt.
, nagos, radzenē un acs lēcā, starp dažu orgānu (piemēram, aknu) intersticiālajām telpām utt.
Tā struktūra piešķir tai īpašas mehāniskās iespējas; tai ir liela mehāniskā izturība, kas saistīta ar augstu elastību (piemēram, cīpslās) vai augstu stingrību (piemēram, kaulos) atkarībā no funkcijas, kas tai jāveic.
Viena no interesantākajām kolagēna īpašībām ir tās vienkāršā vienkāršība: to veido aptuveni 30% prolīna un aptuveni 30% glicīna; pārējām 18 aminoskābēm jāsadala atlikušie 40% olbaltumvielu struktūras. Kolagēna aminoskābju secība ir ārkārtīgi regulāra: uz katriem trim atlikumiem trešā ir glicīns.
Prolīns ir cikliska aminoskābe, kurā R grupa saistās ar α-amino slāpekli, un tas piešķir tai zināmu stīvumu.
Galīgā struktūra ir atkārtota ķēde ar spirāles formu; kolagēna ķēdē ūdeņraža saites nav. Kolagēns ir "kreisās puses spirāle, kuras solis (garums atbilst vienam spirāles pagriezienam) ir lielāks nekā" α-spirāle; kolagēna spirāle ir tik vaļīga, ka trīs olbaltumvielu ķēdes spēj aptīt viena otru, veidojot " viena virve: trīskāršas spirāles struktūra.
Trīskāršā kolagēna spirāle tomēr ir mazāk stabila nekā gan α-spirāle, gan β-loksnes struktūra.
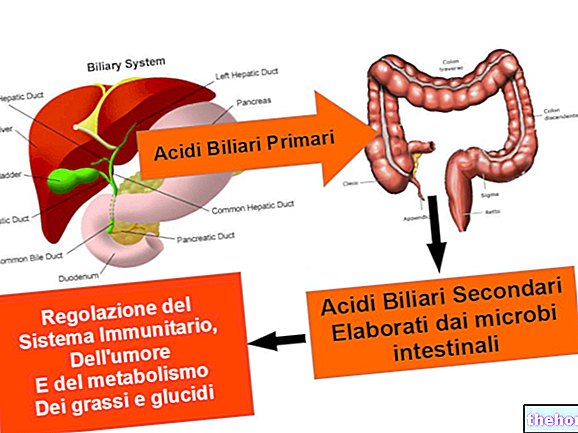
Tagad apskatīsim mehānismu, ar kuru tiek ražots kolagēns; ņemiet vērā, piemēram, asinsvadu plīsumu: šo plīsumu pavada neskaitāmi signāli ar mērķi aizvērt trauku, tādējādi veidojot recekli.
Koagulācijai nepieciešami vismaz trīsdesmit specializēti fermenti. Pēc recekļa ir jāturpina audu remonts; šūnas, kas atrodas tuvu brūcei, ražo arī kolagēnu. Lai to izdarītu, vispirms tiek ierosināta gēna ekspresija, tas ir, organismi, kas, sākot ar gēna informāciju, spēj ražot proteīnu (ģenētiskā informācija tiek pārrakstīta uz mRNS, kas atstāj kodolu un sasniedz ribosomas citoplazmā, kur ģenētiskā informācija tiek pārvērsta proteīnā.) Tad kolagēns tiek sintezēts ribosomās (izskatās kā kreisās puses spirāle, kas sastāv no aptuveni 1200 aminoskābēm un kuras molekulmasa ir aptuveni 150 000 d), un pēc tam uzkrājas lūmenos, kur tas kļūst par fermentu substrātu, kas spēj veikt pēctranslācijas modifikācijas (valodas izmaiņas, kas tulkotas ar "mRNS"); kolagēnā šīs izmaiņas sastāv no dažu sānu ķēžu, īpaši prolīna un lizīna, hidroksilēšanas.
Fermentu neveiksme, kas noved pie šīm izmaiņām, izraisa skorbutu: tā ir slimība, kas sākotnēji izraisa asinsvadu plīsumu, zobu lūzumu, kam var sekot starpzarnu asiņošana un nāve; to var izraisīt nepārtraukta ilgstošas lietošanas pārtikas lietošana.
Pēc tam citu fermentu darbības dēļ notiek citas izmaiņas, kas ietver prolīna un lizīna hidroksilgrupu glikozīdēšanu (cukurs saistās ar OH skābekli); šie fermenti ir atrodami citos apgabalos, nevis lūmenā, tāpēc proteīns, veicot izmaiņas, migrē endoplazmatiskajā retikulā, lai nonāktu maisiņos (pūslīšos), kas aizveras un atdalās no tīklojuma: to iekšpusē ir glikozidēts pro -kolagēna monomērs; pēdējais sasniedz Golgi aparātu, kur konkrēti fermenti atpazīst cisteīnu, kas atrodas glikozīdētā pro-kolagēna karboksiālā gala daļā, un izraisa dažādu ķēžu tuvināšanos viena otrai un veido disulfīda tiltus: šādā veidā trīs ķēdes glikozīdētais pro-kolagēns tiek iegūts savstarpēji saistīts, un tas ir sākumpunkts, no kura trīs ķēdes, savstarpēji caurplūstot, spontāni rada trīskāršu spirāli. Trīs savstarpēji saistītās glikoksidētā pro-kolagēna ķēdes sasniedz vezikulu, aizrijoties ar sevi, atdalās no Golgi aparāta, transportējot trīs ķēdes uz šūnas perifēriju, kur caur kausējumu jonu ar plazmas membrānu, trimetrs tiek izvadīts no šūnas.
Papildu šūnu telpā ir īpaši fermenti, pro-kolagēna peptidāzes, kas no šūnas izņemtajām sugām noņem trīs fragmentus (viens katrai spirālei) no 300 aminoskābēm 1 ", no gala karboksīda daļas un trīs fragmenti (pa vienam katrai spirālei), kuros katrā ir aptuveni 100 aminoskābes, no aminoterminālās daļas: paliek trīskāršā spirāle, kas sastāv no aptuveni 800 aminoskābēm vienā spirālē, kas pazīstama kā tropokollagēns.
Tropokollagēnam ir diezgan ciets stienis; dažādi trimmeri ir saistīti ar kovalentām saitēm, lai iegūtu lielākas struktūras: mikrošķiedras. Mikrofibrilās dažādi trimmeri ir sakārtoti pakāpeniski; daudzas mikrošķiedras veido tropokologēna saišķus.
Kaulos, starp kolagēna šķiedrām, ir starpnozaru telpas, kurās nogulsnējas kalcija un magnija sulfāti un fosfāti: šie sāļi aptver arī visas šķiedras; tas padara kaulus stīvus.
Cīpslās starpnozaru telpās ir mazāk kristālu nekā kaulos, savukārt olbaltumvielas ir mazākas nekā tropokologēnā: tas piešķir cīpslām elastību.
Osteoporoze ir slimība, ko izraisa kalcija un magnija trūkums, kā dēļ nav iespējams fiksēt sāļus tropokologēna šķiedru intersticiālajās zonās.