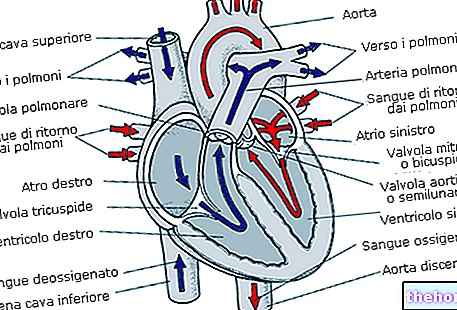
Glikogēns ir α-glikozes makromolekula (molekulmasa aptuveni 400 miljoni daltonu), kurā galvenokārt ir α-1,4 glikozīdiskās saites un sekas 1:10, pateicoties α-1,6 glikozīdu saitēm.
Glikogēns ir rezerves materiāls un tiek nepārtraukti sadalīts un atjaunots; visā ķermeņa šūnu masā ir aptuveni 100 g glikogēna: lielākā daļa no tā atrodas aknās, kur tas ir kustīgs, un tāpēc to var izmantot kā rezervi citiem orgāniem (glikogēns muskuļos nav kustīgs).
Visi fermenti, kas katalizē glikogēna noārdīšanos un sintēzi, atrodas citoplazmā, tāpēc ir nepieciešama regulēšanas sistēma, kas padara vienu ceļu neaktīvu, kad otrs ir aktīvs: ja ir pieejama glikoze, pēdējais tiek pārvērsts par glikogēnu (anabolisms). rezerve, otrādi, ja glikozei ir nepieciešams c ", tad glikogēns tiek noārdīts (katabolisms).

Ferments, kas galvenokārt iesaistīts glikogēna sadalīšanā, ir glikogēna fosforilāze; šis enzīms spēj šķelt glikozīdisko α-1,4 saiti, izmantojot neorganisku ortofosfātu kā lītisku līdzekli: šķelšanās notiek fosforolītiskā veidā un tiek iegūts glikozes 1-fosfāts.
Piecās vai sešās vienībās no sazarošanās punkta glikogēna fosforilāzes enzīms vairs nespēj darboties, tāpēc tas atdalās no glikogēna un tiek aizstāts ar deramificējošu enzīmu, kas ir transferāze: šī fermenta katalītiskajā vietā c "ir" histidīns, kas ļauj pārnest trīs saharīdu vienības uz tuvāko glikozīdu ķēdi (histidīns uzbrūk pirmajam glikozes molekulas ogleklim). Tikko minētais enzīms ir glikoziltransferāze; šī fermenta darbības beigās sānu ķēdē paliek tikai viena glikozes vienība, un pirmais ogleklis ir saistīts ar sesto oglekli no galvenās ķēdes glikozes. Pēdējā glikozes vienība sānu ķēdē tiek atbrīvota, darbojoties "enzīms α-1,6 glikozidāze (šis enzīms veido deramificējošā enzīma otro daļu); ņemot vērā, ka glikogēna zari ir proporcijā 1:10, no pilnīgas makromolekulas noārdīšanās mēs iegūstam aptuveni 90% glikozes 1-fosfāta un aptuveni 10% glikoze.
Iepriekš minēto fermentu darbība ļauj no glikogēna molekulas izņemt sānu ķēdi; šo enzīmu darbību var atkārtot, līdz notiek pilnīga ķēdes noārdīšanās.
Aplūkosim hepatocītu; glikoze (asimilēta ar uzturu), kad tā nonāk šūnā, tiek pārvērsta par glikozes 6-fosfātu un tādējādi tiek aktivizēta. Glikozes 6-fosfāts, iedarbojoties fosfoglukomutaze, tiek pārveidots par glikozes 1-fosfātu: pēdējais nav tiešs biosintēzes priekštecis; biosintēzē tiek izmantota aktivēta cukura forma, ko attēlo cukurs, kas saistīts ar difosfātu: parasti uridildifosfāts (UDP). Glikozes 1-fosfāts ir pēc tam pārvēršas par UDP-glikozi, šo metabolītu iedarbībā glikogēna sintāze kas spēj saistīt UDP-glikozi ar augošā glikogēna nesamazināmo galu: tiek iegūts glikozīdās vienības iegarenais glikogēns un UDP.
Glikogēna noārdīšanās notiek, iedarbojoties glikogēna fosforilāze kas izdala glikozes molekulu un pārveido to par glikozes 1-fosfātu. Pēc tam fosfoglukomutaze glikozes 1-fosfātu pārvērš par glikozes 6-fosfātu.
Glikogēns galvenokārt tiek sintezēts aknās un muskuļos: organismā ir 1–1,2 hektogramma glikogēna, kas sadalīts visā muskuļu masā.
Miocītu glikogēns veido enerģijas rezervi tikai šai šūnai, savukārt aknās esošais glikogēns ir rezerves avots arī citiem audiem, tas ir, to kā glikozi var nosūtīt uz citām šūnām.
Glikozes 6-fosfāts, kas iegūts muskuļos glikogēna noārdīšanās rezultātā, enerģijas pieprasījuma gadījumā tiek nosūtīts uz glikolīzi; aknās glikozes 6-fosfāts tiek pārveidots par glikozi, iedarbojoties uz glikozes 6-fosfāta fosfatāze (raksturīgs hepatocītu enzīms) un tiek nogādāts asinsritē.
Glikogēna sintāze un glikogēna fosforilāze darbojas uz nesamazinošām glikogēna vienībām, tāpēc jābūt hormonālam signālam, kas pavada viena ceļa aktivizēšanu un otra bloķēšanu (vai otrādi).
Laboratorijā bija iespējams pagarināt glikogēna ķēdi, izmantojot glikogēna fosforilāzi un izmantojot ļoti augstu glikozes 1-fosfātu.
Šūnās glikogēna fosforilāze katalizē tikai noārdīšanās reakciju, jo metabolītu koncentrācija ir tāda, kas novirza šādas reakcijas līdzsvaru uz labo pusi (ti, uz glikogēna noārdīšanos):

Apskatīsim glikogēna fosforilāzes darbības mehānismu: acetāla skābeklis (kas darbojas kā tilts starp glikozes vienībām) saistās ar fosforila ūdeņradi: veidojas reakcijas starpprodukts, ko piešķir karbokācija (uz glikozes, kas ir viss "). ekstremitātes), pie kurām fosforils (Pi) saistās ļoti ātri.
Glikogēna fosforilāzei ir nepieciešams kofaktors, kas ir piridoksāla fosfāts (šī molekula ir arī transamināžu kofaktors): tai ir daļēji protonēts fosforils (piridoksāla fosfātu ieskauj hidrofobiska vide, kas attaisno ar to saistīto protonu klātbūtni). Fosforils (Pi) spēj pārnest protonu uz glikogēnu, jo šis fosforils pēc tam atkārtoti iegūst protonu no piridoksāla fosfāta daļēji protonētā fosforila. Varbūtība, ka pie fizioloģiskā pH fosforils zaudē protonu un paliek pilnīgi deprotonēts, ir ļoti zema.
Tagad redzēsim, kā darbojas fosfoglukomutaze.Šis enzīms katalītiskajā vietā uzrāda fosforilēta serīna atlikumu; serīns iegūst fosforilu par glikozes 1-fosfātu (sestajā pozīcijā): īsu laiku veidojas 1,6-bisfosfāts glikoze, pēc tam serīns tiek pārfosforilēts, ieņemot fosforilu pirmajā pozīcijā. Fosfogluko mutāze var darboties abos virzienos, ti, pārvēršot glikozes 1-fosfātu par glikozes 6-fosfātu vai otrādi; ja tiek ražots glikozes 6-fosfāts, to var nosūtīt tieši uz glikolīzi muskuļos vai pārveidot par glikozi aknās.
Ferments uridilfosfogliko transferāze (vai UDP glikozes pirofosforilāze) katalizē glikozes 1-fosfāta pārnešanas reakciju uz UTP, pievienojoties fosforilam a.
Tikko aprakstītais enzīms ir pirofosforilāze: šis nosaukums ir saistīts ar faktu, ka pretējā reakcija pret tikko aprakstīto ir pirofosforilēšana.
UDP glikoze, kas iegūta, kā aprakstīts, spēj pagarināt glikogēna ķēdi, izmantojot monosaharīdu vienību.
Ir iespējams panākt, lai reakcija attīstītos uz UDP glikozes veidošanos, likvidējot produktu, kas ir pirofosfāts; fermentu pirofosfatāze pārvērš pirofosfātu divās ortofosfāta molekulās (anhidrīda hidrolīze) un, to darot, saglabā pirofosfāta koncentrāciju tik zemu, lai UDP glikozes veidošanās process būtu termodinamiski labvēlīgs.
Kā jau minēts, UDP glikoze, pateicoties glikogēna sintāzes darbībai, spēj pagarināt glikogēna ķēdi.
Sekas (proporcijā 1:10) ir saistītas ar faktu, ka tad, kad glikogēna ķēde sastāv no 20–25 vienībām, tiek iejaukts sazarojošs enzīms (kura katalītiskajā vietā ir “histidīns”), kas spēj pārnest virkni 7-8 glikozīdu vienības tālāk no 5-6 vienībām: tādējādi tiek radīts jauns sazarojums.
Nervu izcelsmes iemeslu dēļ vai ja enerģija ir nepieciešama fiziskas piepūles dēļ, adrenalīns tiek izdalīts no virsnieru dziedzeriem.
Adrenalīna (un noradrenalīna) mērķa šūnas ir aknas, muskuļi un taukaudi (pēdējās notiek triglicerīdu noārdīšanās un taukskābju cirkulācija: līdz ar to glikoze tiek ražota mitohondriju 6 -fosfātā). tiek nosūtīts uz glikolīzi, bet adipocītos glikozes 6-fosfāts tiek pārveidots par glikozi, iedarbojoties uz glikozes 6-fosfāta fosfatāzi un tiek eksportēts uz audiem).
Redzēsim, tagad adrenalīna darbības metodes. Adrenalīns saistās ar receptoru, kas novietots uz šūnu membrānas (no miocītiem un hepatocītiem), un tas nosaka signāla tulkojumu no ārpuses uz šūnas iekšpusi. Tiek aktivizēta proteīna kināze, kas vienlaicīgi iedarbojas uz sistēmām, kas regulē glikogēna sintēzi un noārdīšanos:
Glikogēna sintāze pastāv divās formās: defosforilēta (aktīva) forma un fosforilēta (neaktīva) forma; proteīnkināze fosforilē glikogēna sintāzi un bloķē tās darbību.
Glikogēna fosforilāze var pastāvēt divos veidos: aktīvā formā, kurā ir fosforilēts serīns, un neaktīvā formā, kurā serīns tiek defosforilēts. Glikogēna fosforilāzi var aktivizēt enzīms glikogēna fosforilāzes kināze. Glikogēna fosforilāzes kināze ir aktīva, ja tā ir fosforilēta, un neaktīva, ja tā tiek defosforilēta; proteīna kināzes substrāts ir glikogēna fosforilāzes kināze, tas ir, tā spēj fosforilēt (un līdz ar to aktivizēt) pēdējo, kas savukārt aktivizē glikogēna fosforilāzi.
Kad adrenalīna signāls ir beidzies, ir jābeidzas arī tā ietekmei uz šūnu: fosfatāzes enzīmi iejaucas olbaltumvielu sugās.
.jpg)